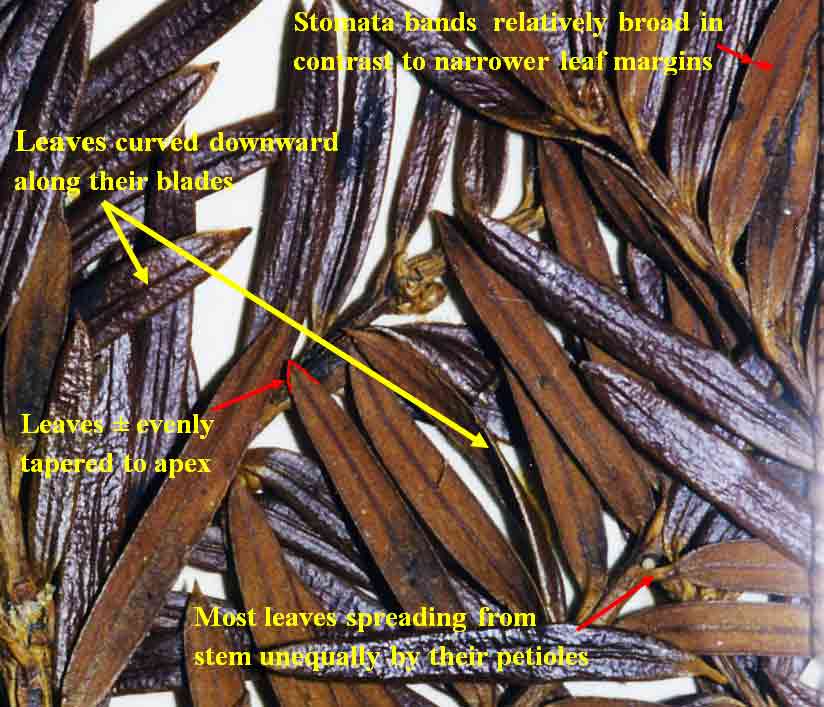
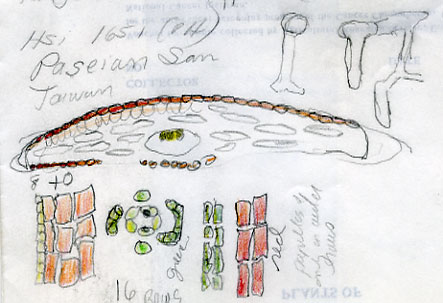
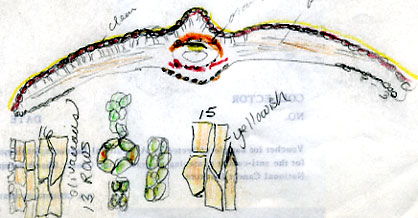
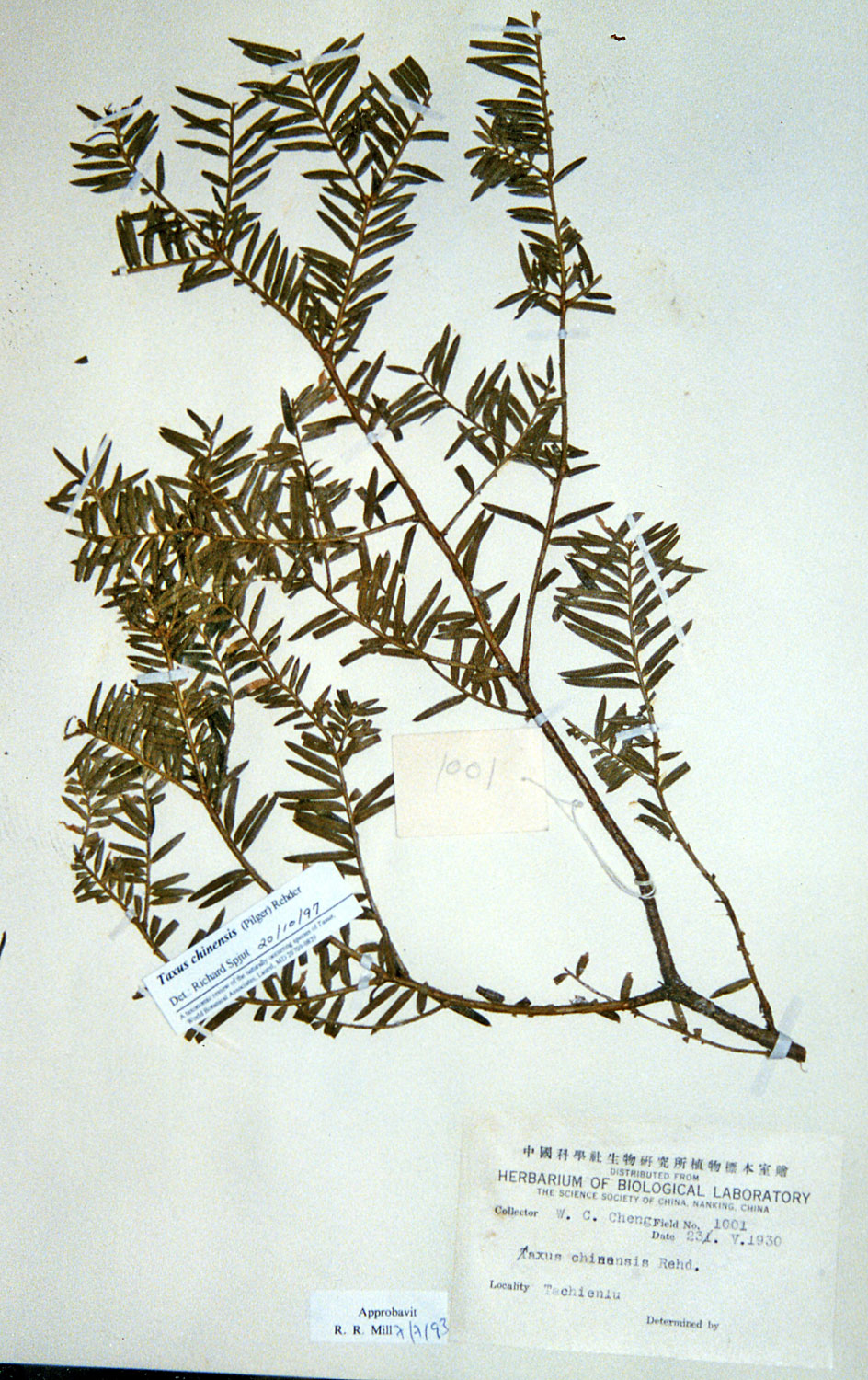
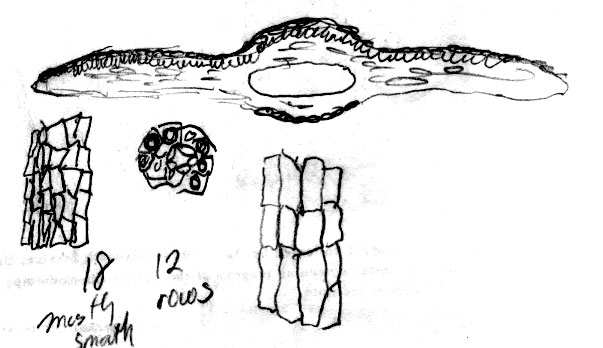
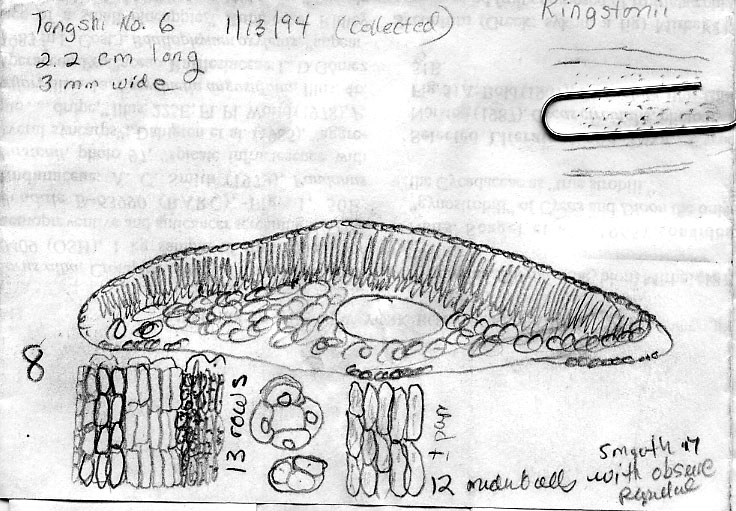
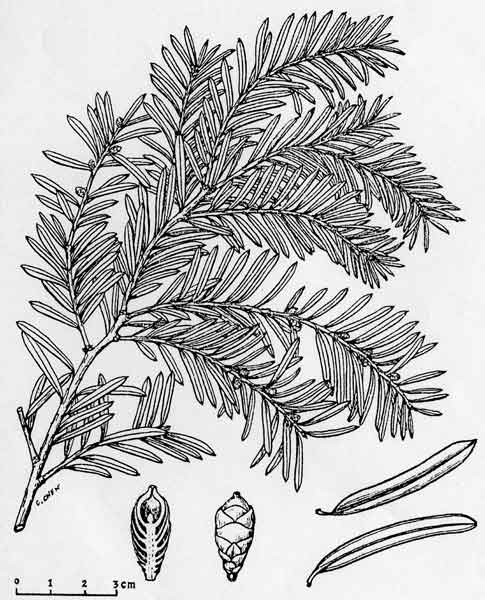
13. Taxus kingstonii Spjut , J. Bot. Res. Inst. Texas 1(1): 240. 2007. Type: China. Taiwan: Arisan Prov., Kagi, 2833 m [Mt. Alishan], 2 Feb 1918, tree 25 ft. x 2 ft., only one seen, Wilson 9738, holotype: A! (with male cones; leaf with 10–11 stomata rows/band, abaxial margin with 8 smooth, thick-walled, trapezoidal cells across, anticlinal to periclinal in arrangement, followed by 5 rows of papillose cells, midrib 12 cells wide, mostly smooth, papillose on outer 2 rows); isotypes: BM! (leaf with 13 stomata rows and 9 marginal smooth cells, midrib mostly smooth except upper third of leaf, papillose on outer 2 rows of cells), K! US (p.p.)! Kingston yew. Forest margins, or summits, 2450-2833 m in Taiwan, to 800 m on mainland Asia; India (Khasi Hills), Myanmar, China (Tibet, Gansu, Shaanxi, Sichuan, Yunnan, Taiwan). Taxus kingstonii is recognized by the relatively turgid recurved leaves (twist and curve downwards along their blades) that taper rather evenly to apex, and by the rusty orange color on the abaxial leaf surface (dried specimens). In herbarium specimens, the leaves often crisscross, especially near apex of branchlets, whereas leaves in other species of the Sumatrana Group—that are reddish or greenish in color—appear more evenly two-ranked and tightly adpressed to branchlets. These differences seem related to the manner in which leaves twist, which may also correlate with leaf shape as seen by the evenly tapered leaf in the Kingston yew compared to the falcate leaf in other species (indented more along lower margin near petiole; Fig. 20, 21, 30, 31). Other features that help distinguish the Kingston yew are seen on the abaxial leaf midrib (note: 27 June 2006—as seen mostly from specimens from Khasia, Tibet, Gansu, Sichuan, and Taiwan, but not in some specimens from Yunnan). It usually forms a rounded keel, in contrast to a truncated ridge that may either be flush with the surface in T. celebica, or elevated in T. mairei. Its epidermal cells are short trapezoidal to almost rectangular, and in T-section appear similar in size and shape to those on the adaxial surface, or the adaxial cells sometimes are larger and nearly isodiametric. Taxus mairei, on the other hand, has shorter trapezoidal cells on its abaxial midrib, but in T-section they appear larger and nearly globose—in contrast with the shorter elliptical cells on the adaxial surface. The abaxial midribs of the related T. sumatrana and T. celebica have longer, nearly rectangular cells that in T-section are similar in size on both surfaces. The Kingston yew also has conspicuous persistent bud-scales at the base of young branchlets, in contrast to those of related species in the Sumatrana Group that appear vestigial as in T. chinensis. The Kingston yew generally occurs at elevations between that of T. mairei (below 1200 m) and that of T. wallichiana (above 2300 m). Variation ascribed to T. kingstonii may include hybrids with T. chinensis and T. mairei in Shaanzi, Gansu and Sichuan, T. celebica in Yunnan and Burma, and T. wallichiana in NE India. These alleged hybrids, however, show disjunct geographical relationships as further described in the examples that follow. (A) Four collections, two from Shaanxi, one from Gansu, and another from Sichuan that I had annotated T. chinensis without microscopic examination of leaves, were later found to lack papillae on the abaxial midrib, a feature that I employ to distinguish T. chinensis from T. kingstonii. Their leaves spread nearly at right angles—parallel to each other—and have the greenish color of T. chinensis; otherwise, they compare favorably with typical T. kingstonii. The close morphological similarity among these specimens, along with their more northern geographical occurrence, suggests a distinct taxon. (B) Two collections, one from Taiwan (Liu et al. 389) and one from Yunnan (Forrest 12087), resemble T. mairei by the isodichotomous branching, leaves spreading at right angles, and by the thin-walled leaf epidermal cells with a yellowish chloroplast, but are regarded T. kingstonii by the rusty orange color on leaves and branchlets, and by the leaves ± evenly tapered to base and apex. The abaxial leaf epidermal cells in the specimen from Yunnan are not enlarged on the midrib—further supporting its determination as T. kingstonii, whereas the Taiwan specimen has slightly larger cells—indicating that it could be assigned to T. mairei. Yet, the closer similarity between these specimens than to other specimens indicates a common but distinct ancestry. On the other hand a more conservative treatment might include the Philippine yews (T. obscura ined.) under T. kingstonii based on leaf arrangement and color, a character that I had applied to a limited extent that resulted in other Philippine specimens being included under T. kingstonii in my 1996 annotations (e.g., Leano 25128). This, however, seems contrary to leaf anatomical data in this study. The relatively narrow border of bare marginal cells (4 cells wide), the papillose midrib, and the 11–14 stomata rows/band occur consistently in many specimens that would seem to merit separate taxonomic status. It is also interesting that Ludlow & Sherriff 3719 from NE India has similarly discolored leaf surfaces with elliptical shaped epidermal cells in T-section, 12 stomata rows/band, and a papillose undersurface that extends entirely across the reddish marginal zone, and that Tsai 59874 from Yunnan, and that Neth. Ind. For. Serv. bb:20887 from Sulawesi, are also similar in this regard. Their leaf characteristics compare more with The Philippines yews than with those in the Himalayas. Further study is needed to determine to what extent epidermal papillae are influenced by genetic and environmental factors. Taxus kingstonii is named in honor of David G. I. Kingston, a chemist who has done extensive work on elucidating and summarizing the taxane chemistry of the genus (e.g., Kingston 1996; Kingston et al. 1990. Taxol, from which the drug paclitaxel is marketed and used to treat ovarian and other cancers, was originally isolated from dried bark of T. brevifolia; however, fresh leaves of many species of Taxus have proved suitable for obtaining taxanes. Taxol was first characterized by Monroe Wall’s group (Research Triangle Institute, Wani et al. 1971). They also discovered other significant antitumor agents such as camptothecin and holacanthone (Wall et al. 1972). Representative Specimens—India: Khasia, Hooker L77 (PH), 1337 (K) Simmons 484 (P), Nungluai, 5000 ft, Mann (K, P). Myanmar: Bernardmyo, Ruby Mines, 5600 ft (K). China—Tibet: Oriental, Haut Mekong, Tsekou to Nekou, Soulie1411 (P: 2 sheets). Gansu: F. N. Meyer 1790 ex USDA (P). Shaanxi (Shensi) Davis 1872 (P). Tsin-lin au Lao-lin, 3000 m. Sichuan: Tachienlu Cheng 1001 (BM), Cheng 1475 (P). Yunnan: Shweli-Salween Divide, 10,000 ft, 25º20 N, shrub 10–20 ft, open shady thickets, Forrest 11789 (BM, K), Salween, Forrest 12087 (K, S: C-2093), Forrest s.n. (A); Ma-Chang-Kai, valley, 25º30 N, 6000 ft., shrub, 20–30 ft., in thickets, Dec 1918, Forrest 9462 (A, K), Forrest 15945 (BM, K); Salween E of Tengyueh, to summit of Shwell, Shwelli River, Rock 7587 (US). Taiwan: Paseian San [Pahsienshan], Hsi 165 (PH); Fig. 3 in Li, Woody Fl. Taiwan, Liu et al. 437, (M, T, US); Mt. Ammachan, Liu 0389 (A, K), Arizan, Nitak (PH); Mt. Ammashan, Taichung Hsien, C.C. Tseng s.n. (BH); Tongshi, C-j. Chang, Dongshi #2, Tongshi #6, without locality data, 4 Mar 1993 (wba).
|
||||

|
Taiwan: Left: Taxus kingstonii isotype
(
|
Yunnan: Left—Forrest 9339 (BM) and close-up of the same. Right two photos—Forrest s.n. (A) and close-up. These specimens have scales at base of branchlets that are similar to T. wallichiana. The leaves on the abaxial surface show an elevated-channeled midrib and glossy margin, which was further confirmed in microscopic examination as shown in the illustrations on left below. These features suggest T. sumatrana except that the leaves are not puckered. The rusty orange coloration, tapered leaf bases, and leaves evenly tapered to apex favor T. kingstonii. The two lower photos on right also specimens collected by Forrest from Yunnan (Forrest 11789, K); scales and seed can be seen in the lower right photo.
Khasia: Left two photos, Hooker 77 (P), showing specimen and close-up of seed and scale at base of branchlet; right two photos, Hooker 1337 (K), specimen and illustration. The illustration indicates epidermal cells under the microscope appear reddish-orange on adaxial surface, yellowish on abaxial surface, clear in the the palisade parenchyma, and greenish in the stomata bands. These specimens hardly differ from those collected from Yunnan as shown above.
|
  
Gansu:
|
Yunnan: |
|
|
Note that leaves on the adaxial surface are strongly convex in contrast to a more level surface in T. mairei. |
|
|
|
|
|