|
Niebla testudinaria is a fruticose lichen
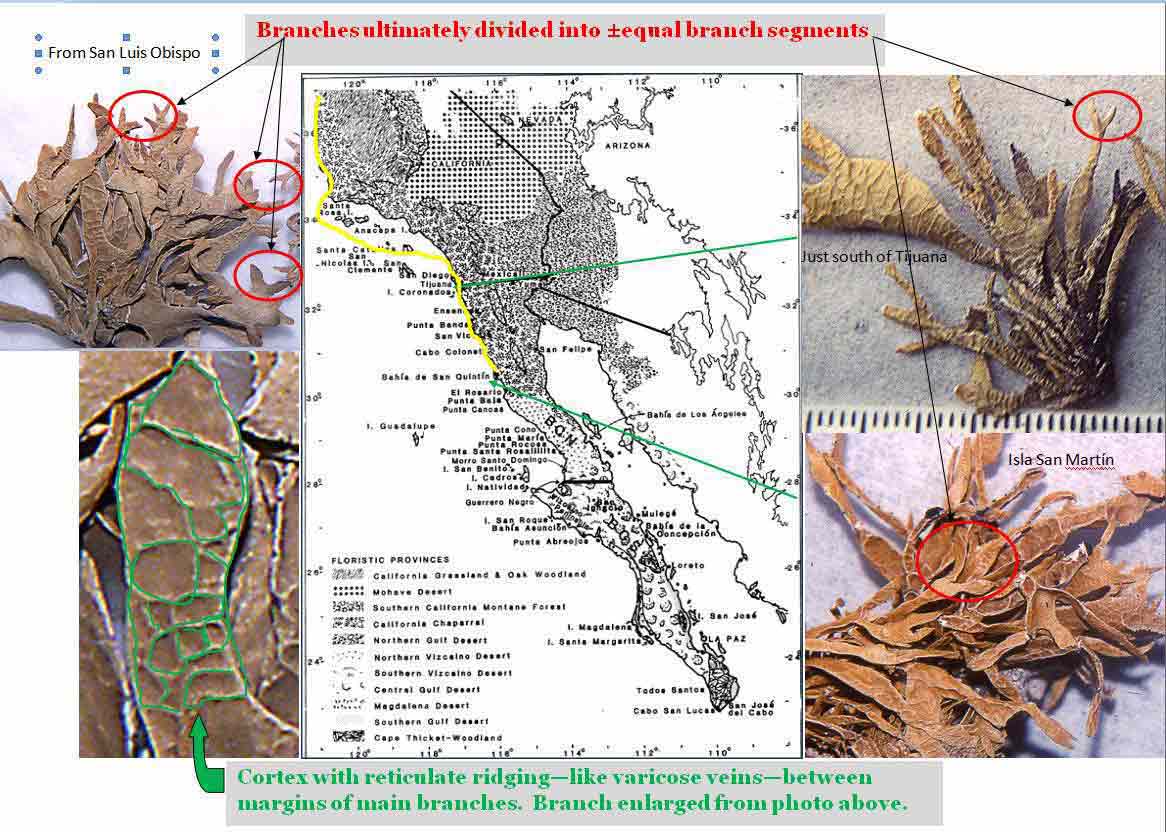
found on rocks along the Pacific coast from Marin County, California to
near Punta Santa Rosalillita in Baja California, and also in the Channel Islands
south into Mexico on Isla San Martín (just off the
coast of San Quintín), and west to Isla Guadalupe. It
appears to have been most common on the Monterrey Peninsula such as in Point Lobos State Park,
where also occurring with
Niebla eburnea,
Vermilacinia procera and V. laevigata.
During the mid to late 1980's, it was observed to be
locally common just south of Tijuana near Rosarito on
talus-like rubble in association with Vermilacinia
combeoides and V. pumila. This area has since undergone extensive real estate development.
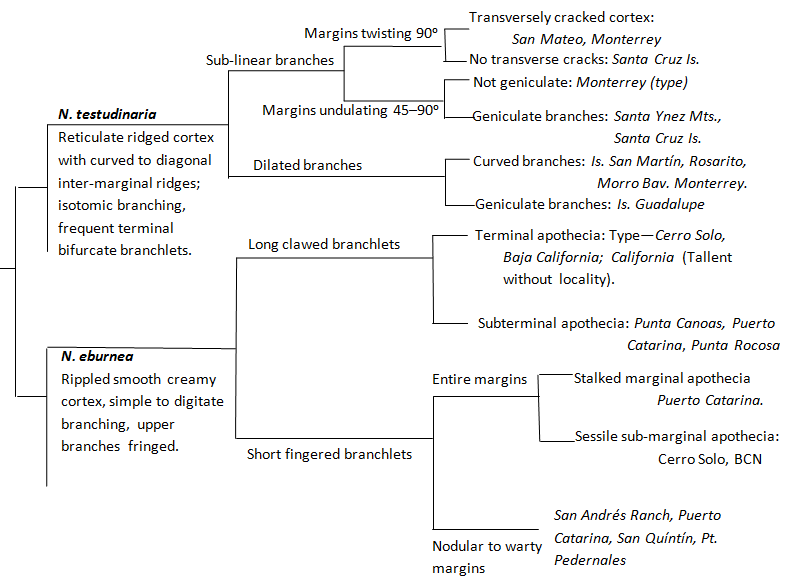
Niebla testudinaria is recognized by the primary branches spreading
outwards from their base, in contrast to the upright habit of the fastigiate
branches that characterizes
Niebla homalea, and by reticulate cortical
ridges on the face of the branches in contrast to the relatively smooth surface
of Niebla eburnea.
The epithet, testudinaria, provided by Nylander, suggests that
the species was recognized for the reticulate pattern on the cortical surface,
like that seen on tortoise shells, especially thalli with
dilated branch segments.
The varicose vein-like ribbing within a relatively thick cortex (>75
µm), along with the presence of divaricatic acid, also distinguishes N. testudinaria from
related species;
Niebla homalea, for example, has relatively
narrow linear branches with a glossier cortex transversely
cracked at various intervals. Another characteristic feature of N. testudinaria is that
branches terminally divide
± equally,
dichotomous or digitately, especially those that appear arcuate and compressed
where they divide. This is in contrast to unequal
divisions of the primary linear subterete branches of
N.
homalea. The
reticulate ridging that characterizes Niebla testudinaria is seen in thalli with narrow
to irregularly widened
branches. The narrower parts appear prismatic in cross-section, the wider parts
often expand and flattened
like the palm of a hand from which
±
isodichtomous or isotrichotomous (isotomic) branchlets arise.
Relatively short thalli, wider
than tall (e.g., Moran 1055 from Isla San
Martín, Leavitt et al. 16-1166)
resemble N. caespitosa, which differs by its relatively thinner
cortex as evident by more sharply defined cortical ridges, and by irregularly
lacerated branches.
Charis Bratt 2303 from Santa Cruz Island (N. caespitosa),
which might be referred to N.
testudinaria, was distinguished by its thinner cortex; otherwise, it is
similar to Spjut & Sérusiaux 17114, and Leavitt et al. 16-829, 16-703
shown above.
Charis Bratt 6431 from Santa Cruz Island and Charis Bratt 7202
from the Santa Ynez Mountains shown above closely resemble
Niebla halei by
their linear shaped prismatic branches; N. halei, described from
San Bruno Mountain, differs by its much smaller thallus. These Charis
Bratt specimens may prove to be a distinct species, or perhaps could be referred
to N. halei in further study.
DNA phylogeny of N. testudinaria, as with most other depside species,
reveals it to occur in different subclades, or microclades; however, statistical analyses
(BPP, Stacey) for three specimens indicated only one species. Specimens
collected by Leavitt et al. were not included in Jorna et al. (2021).
The character feature of reticulate
cortical ribbing of Niebla testudinaria appears plesiomorphic to various
subtaxa that might be formally named. This is exemplified in the following
dendrogram.
References:
Bowler, P. A, R. E.
Riefner, Jr., P. W. Rundel, J. Marsh & T.H. Nash, III. 1994. New species
of Niebla (Ramalinaceae) from western North America. Phytologia
77: 23-37.
Bowler, P. A. and
J. Marsh. 2004. Niebla.
Lichen Flora of the Greater Sonoran Desert 2: 368–380.
O’Brien, H. E., J.
Miadlikowska, and F. Lutzoni 2009. Assessing
reproductive isolation in highly diverse communities of the lichen-forming
fungal genus Peltigera. Evolution 63-8: 2076–2086.
Culberson, C. F., W.
L. Culberson & A. Johnson. 1988. Gene flow in lichens. American Journal of
Botany 75: 1135–1139.
DePriest, P. T. 1994.
Variation in the Cladonia chlorophaea Complex II: Ribosomal DNA variation
in a Southern Appalachian population. The Bryologist 97: 117–126.
Lohtander, K., L. Myllys, R. Sundin, M.
Kllersj, and A. Tehler. 1998. The species pair concept in the lichen
Dendrographa leucophaea (Arthoniales): Analyses based on ITS
Sequences. Bryologist 101: 404–411.
Montagne, D.M. 1852.
Diagnoses phycologicae. Ann. Sci. Nat. Sr. 3, 18, 302-319.
Spjut, R. W. 1995. Vermilacinia (Ramalinaceae,
Lecanorales), a new genus of lichens. Pp. 337-351 in Flechten
Follmann; Contr. Lichen. in honor of Gerhard Follmann, F. J. A. Daniels,
M. Schulz & J. Peine, eds., Koeltz Scientific Books, Koenigstein.
_________. 1996. Niebla and Vermilacinia (Ramalinaceae)
from California and Baja California. Sida, Botanical Miscellany 14:
1–207, 11 plates.
Additional References: See
Niebla.
|














{kind=link}
{kind=link}