|
Niebla and Vermilacinia (Ramalinaceae) from California and Baja
California.
Evolutionary history of coastal species
of fog lichen genera
Spjut R, Simon A, Guissard M, Magain N, Sérusiaux E. The
fruticose genera in the Ramalinaceae (Ascomycota, Lecanoromycetes):
their diversity and evolutionary history.
MycoKeys. 2020 Oct
30;74:109-110].
|
||
|
Key to the
genera Niebla and Vermilacinia
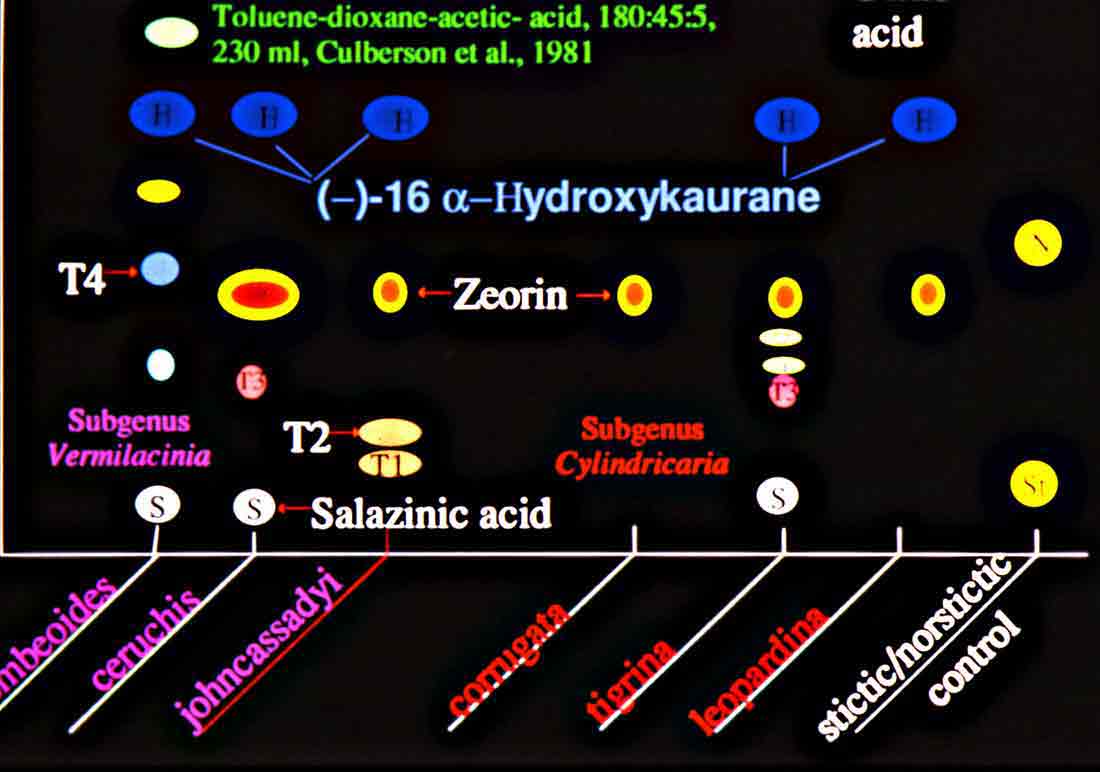
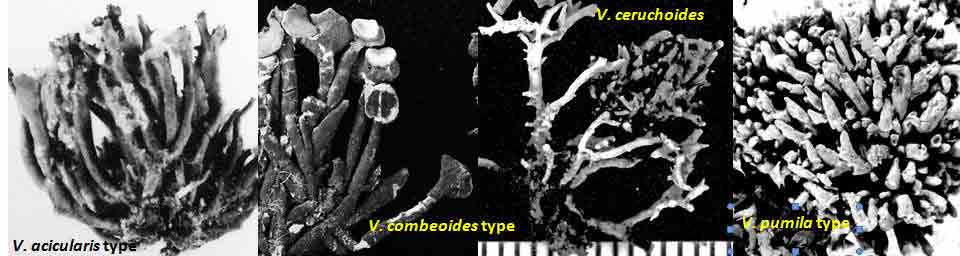
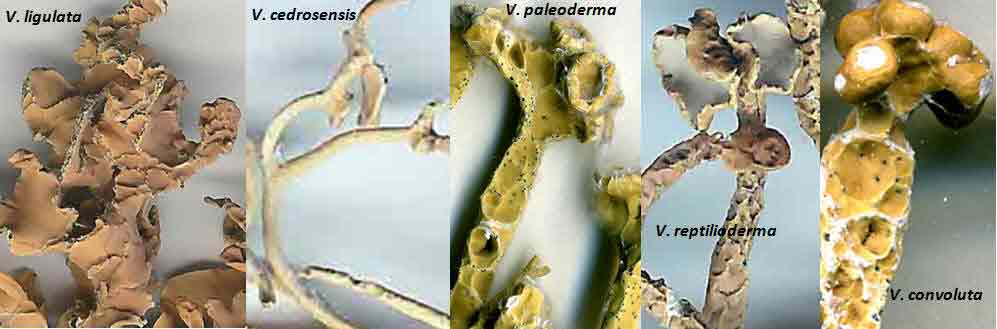
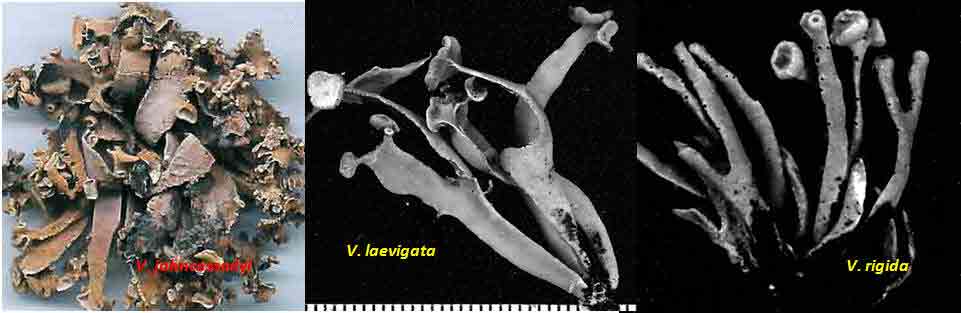
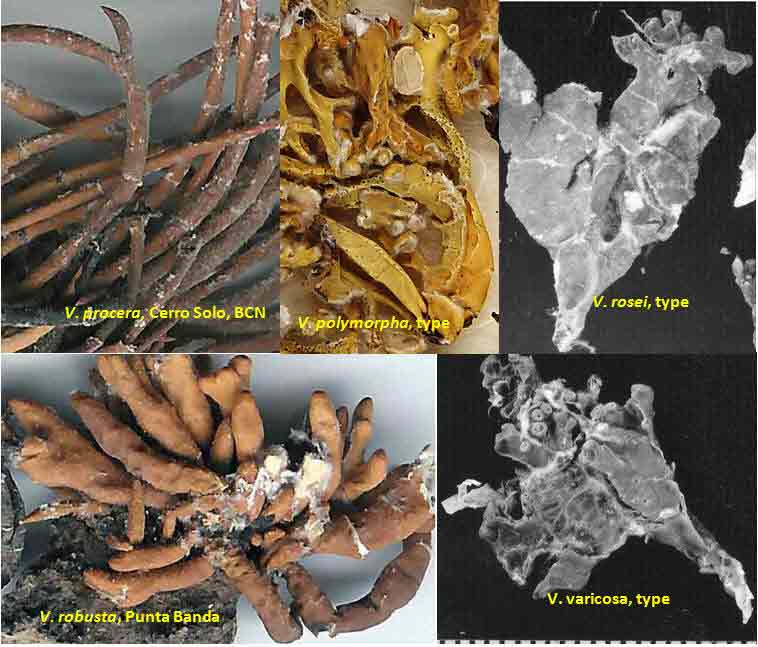
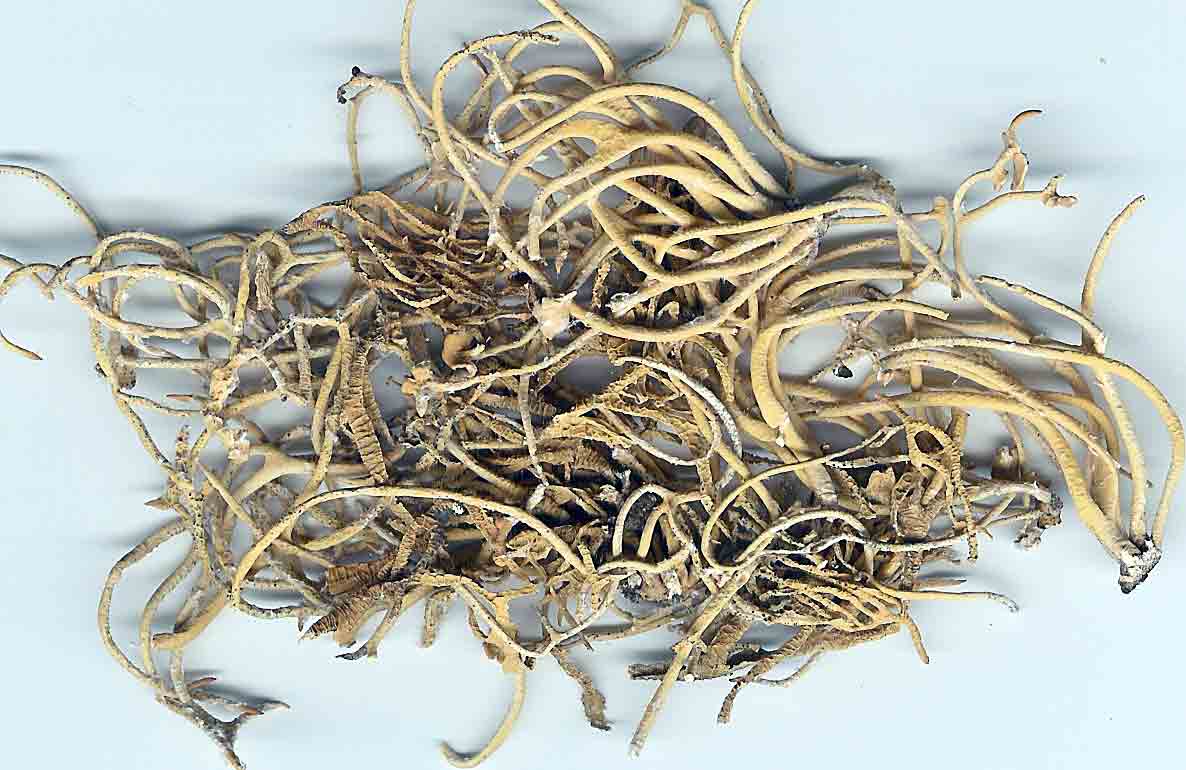
Introduction Vermilacinia is largely a New World genus of 34+ species of fruticose lichens in the family Ramalinaceae characterized by terpenoid secondary metabolites of the diterpene [-] -16 α-hydroxykaurane ('ceruchinol', Lehn and Huneck 1965; Huneck and Follmann 1966; Munoz et al. 2024) and the triterpene zeorin—except for a V. corrugata complex (3 spp.) that lacks ceruchinol—and by the thallus divided into narrow cylindrical (worm-like) branches, either united by a holdfast with erect to pendant branches when growing on shrubs—subgenus Cylindricaria (1o spp., Spjut 1996 + V. granulans, + 2 undescribed in a V. corrugata complex, Spjut et al. 2020), or without a central basal attachment when growing on earth or rock—subgenus Vermilacinia (22+ spp.; Spjut 1996, Spjut et al. 2020). It should be noted that in Huneck and Yoshimura (1996, p. 360), ceruchiol is referred to as (-)-ent-Kauran-16α-ol), and in Rakotondraibe, Spjut & Addo (2024, p. 203, Fig. 16), it is ent-16ß-hydroxykaurane. Key secondary in Vermilacinia have yet to be identified (Rakotondraibe, Spjut & Addo (2024); in Spjut (1996), these include "T1," "T2," etc. Although two subgenera were described (Spjut 1996), a phylogenetic analysis revealed a small sister subgroup within each subgenus, saxicolous V. laevigata and V. combeoides in the corticolous Cylindricaria, and V. procera sister to the other saxicolous Vermilacinia (Spjut et al. (2020). Thus, four subgenera might be recognized for Vermilacinia, however, not all saxicolous species have been sampled for their DNA. The type species for the genus is V. combeoides. Initially 28 species of Vermilacinia were distinguished by Spjut (1996) based strictly on morphology and secondary metabolites, 18 in subgenus Vermilacinia, 10 in subgenus Cylindricaria. Four additional species in Baja California and Baja California Sur were described by Spjut and Sérusiaux supported by phylogenetic analyses of DNA, and they further recognized the epiphytic V. granulans (Sipman 2011) Spjut & Sérus. in the Atacama Desert (Spjut et al. 2020). The species in both subgenera are distributed among the fog deserts of North America, South America and southwestern Africa (Spjut et al. 2020). Most are endemic to the desert region in which they occur. Exceptions are sorediate species, V. zebrina, based on the type from near Arcata, California, found also in Namibia (Spjut et al. 2020), and V. tigrina described by Follmann (1967, Ramalina tigrina) from Cerro Moreno in the Atacama Desert in Chile; V. tigrina differs by having psoromic acid, but recognized by Spjut (1996) to include hypoprotocetraric acid from a collection by Follmann from near Lima, Peru, and to occur in Baja California based on having either salazinic acid or norstictic acid in addition to the typical secondary terpenoid metabolites found in V. ceruchis, not Niebla homalea, which instead has nieblastictanes and nieblaflavicanes (Zhang et al. 2020; Rakotondraibe, Spjut and Addo 2024). Other North American saxicolous species of Vermilacinia that appear to have disjunct occurrences in the Atacama Desert are V. acicularis described from the Channel Islands (Spjut 1996), and V. robusta known mostly from California (Spjut et al. 2020, Vermilacinia key in MycoKeys 74). DNA data for the South American Vermilacinia have yet to be published. Among the corticolous species (subgenus Cylindricaria), a V. corrugata complex of three species endemic to the Baja California peninsula (but only one described)—is characterized by a strongly reticulated cortex and a thallus deficient in ceruchinol. They can be differentiated morphologically (Spjut unpubl.); however, the DNA phylogeny in Spjut et al. (2020) did not include specimens from the type locality. Another group of three species—characterized by having black-bands or irregularly enlarged spots on the cortex and a hydroxykaurane diterpene—show ecogeographic differences on the Baja California peninsula. They include V. howei, V. nylanderi, and V. tigrina that are not fully resolved by DNA. In South America, V. flaccescens differs from most nonsorediate corticolous species by the lichen metabolite tumidulin (methyl 3,5 dichlorolecanorate), which has also been reported in the epiphytic-sorediate V. granulans and the saxicolous V. lacunosa . The California sorediate V. cephalota is phylogenetically distinct in Baja California, and another sorediate species in Baja California Sur (BCS), V. leonis, also in South America (Spjut 1996), was not collected by Spjut and Sérusiaux for Spjut et al. (2020). Vermilacinia has a long history of being included in the genus Niebla, a substitute name given by Rundel and Bowler (1978) for the illegitimate Desmazieria (homonym) given by Montagne (1852) when he combined the saxicolous Ramalina homalea in California (Acharius 1810) with the broadly interpreted South American R. ceruchis; its transfer to Ramalina had been made by De Notaris (1846). The South American V. ceruchis had long been misapplied to other species recognized to occur on rocks, cacti, and shrubs in South America and on shrubs, especially in coastal North America as described by Bowler and Marsh (2004) for Niebla ceruchis, treated as a synonym of V. ceruchis by Spjut (1996). Its type is based on Parmelia ceruchis Acharius (1803), a terricolous thallus collected near Lima, Peru (Spjut 1996). Its morphology is similar to that collected by Darwin from Iquique in Chile, described by him on his specimen in the Taylor Herbarium (FH) as "lying without adhesion on bare sand," "2000 to 3000 ft high where clouds often hang." Subsequently, it has been interpreted to include other characteristic features treated herein as variants of V. ceruchis (unresolved). The broad interpretation of V. ceruchis has also been classified in other genera dating back to Acharius (1803), all bearing the epithet ceruchis in Parmelia, Borrera, Usnea, and Ramalina. Although Montagne (1852) had long studied the South American thalli under these generic names, the type for his illegit. Desmazieria has to be based on the North American Ramalina homalea (Acharius 1810). Moreover, Spjut (1995) subsequently recognized that the South American thalli associated with the epithet 'ceruchis' belong to a new genus that he named Vermilacinia. DNA studies have since shown Vermilacinia to be phylogenetically distinct from Ramalina and Niebla, as well as from another newly recognized genus, Nambialina, endemic to southwestern Africa (Spjut et al. 2020). Recently, Niebla, Vermilacinia, and Cenozosia (Atacama Desert) have been shown sister to Namibialina, which this relationship included a second newly described species in the genus Cenozosia (Jung et al. 2023), in contrast to the sister relationship of Cenozosia inanis shown in Kistenich (2018). These genera and Ramalina may also constitute a sister group to the genus Cenozosia in which Ramalinopsis (Hawaii) and Trichoramalina (California, Baja California) are included in the ± cosmopolitan Ramalina (Kistenich et. al. 2018). Although “Sérusiaux et al. (2010) provided the first assessment of the North American Niebla (sensu Rundel and Bowler 1978; Bowler and Marsh 2004),” “sampling was limited to relatively few accessions of Vermilacinia” (Spjut et al. 2020); thus, the two gene phylogeny that reportedly supported Niebla (Sérusiaux et al. (2010) actually supported the genus Vermilacinia (Spjut 1995, 1996), which was not accounted for in Sérusiaux et al. (2010). Vermilacinia morphologically differs from Niebla by the absence of twine-like medullary hyphae referred to as chrondroid strands. In Niebla, multiple chondroid strands develop like optical fibers oriented longitudinally within a network of flexuous single hyphal strands. all surrounded by a double-layered cortex. The strands are often visible to the naked eye in cross section. The overlying cortex of Niebla exhibits various patterns of reticulate ridging that aid in distinguishing its species (e.g., ladder-like, N. rugosa with divaricatic acid, N. siphonoloba with sekikaic acid) in contrast to the irregular development of cortical ridging in saxicolous Vermilacinia. The texture of the cortex in saxicolous species appears more like a pie-like crust and its medulla in thallus sections appears like tissue paper under a dissecting scope. The corticolous species generally have a more flexible thallus organization that relates to swelling and shrinking as moisture is absorbed and lost from the seasonal daily formation and dissipation of fog. Its cortex acts like a pliable membrane overlying an accordion-like network of medullary bundles of hyphae ± periclinally oriented and knotted together at frequent intervals. Vermilacinia further differs in its secondary metabolites of zeorin and/or the diterpenoid [-] -16 α-hydroxykaurane (ceruchinol) in all saxicolous and most epiphytic species with exception to some corticolous species, V. corrugata and V. flaccescens. Bourgeanic acid is often present while its taxonomic value has not been fully assessed. Several species within subgenus Vermilacinia have unique triterpenoid compounds endemic to the coastal Vizcaíno Desert where they also have limited geographical occurrences. For example, V. ligulata occurs only along the eastern coast of Isla Cedros and directly across on the adjacent Baja peninsula from where it extends further north about halfway to Punta Canoas, that of V. rosei is known from three scattered collections, Isla San Roque, near Bahía de Tortugas, and on Isla Cedros, and that of V. reptilioderma is restricted to the western Vizcaíno peninsula and nearby Isla Cedros. They may have evolved in isolation from a more widespread ancestral species complex. In Niebla, the depsidone species group may also have evolved on an island in the Vizcaíno Desert Region that—after connection to the mainland peninsula—later spread north to the California chaparral-desert ecotone and west to the Vizcaíno Desert area and from there to the southern part of Isla Cedros. The species taxonomy for saxicolous Vermilacinia described by Spjut (1996) is supported as previously indicated. The key below has been updated in MycoKeys (Spjut et al. 2020). Since this key is open access publication, the following key remains as it was presented on this website in April 2003 with minor changes and updates to the identifications of the images. The new species include: (1) V. breviloba, a relatively small ± tubular inflated thallus with few basal branches, restricted to a narrow canyon inland from Puerto San Andrés; (2) V. lacunosa, found near Bahía Tortugas, distinct for its crateriform surface and the lichen metabolite tumidulin—previously known only from corticolous species in South America; (3) V. pustulata, similar to V. cedrosensis, except the cortex pustular instead of pitted, appearing infrequent but widely distributed in Baja California and Baja California Sur; and (4) Vermilacinia reticulata, distinguished by ± rectangular cortical depressions, appearing of local occurrence near Punta Eugenia.
Key to Niebla
and Vermilacinia
Medulla with twine-like (chondroid) strands; cortex with reticulate
ridging.....................................................
Niebla Key to Subgenera of Vermilacinia
Cortex crustaceous
(like a pie crust), with minute cracks or large
Cortex membranous, 15-30 μm
thick; usually with a regular network Subgenus Vermilacinia |
||
| quently | ||
|
V. ceruchis |
 |

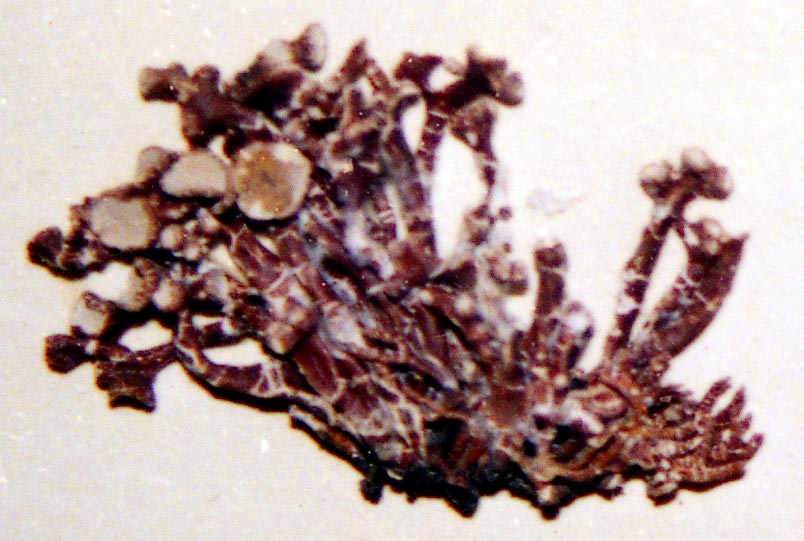
V. ceruchis, typical.
|
 |
||
|
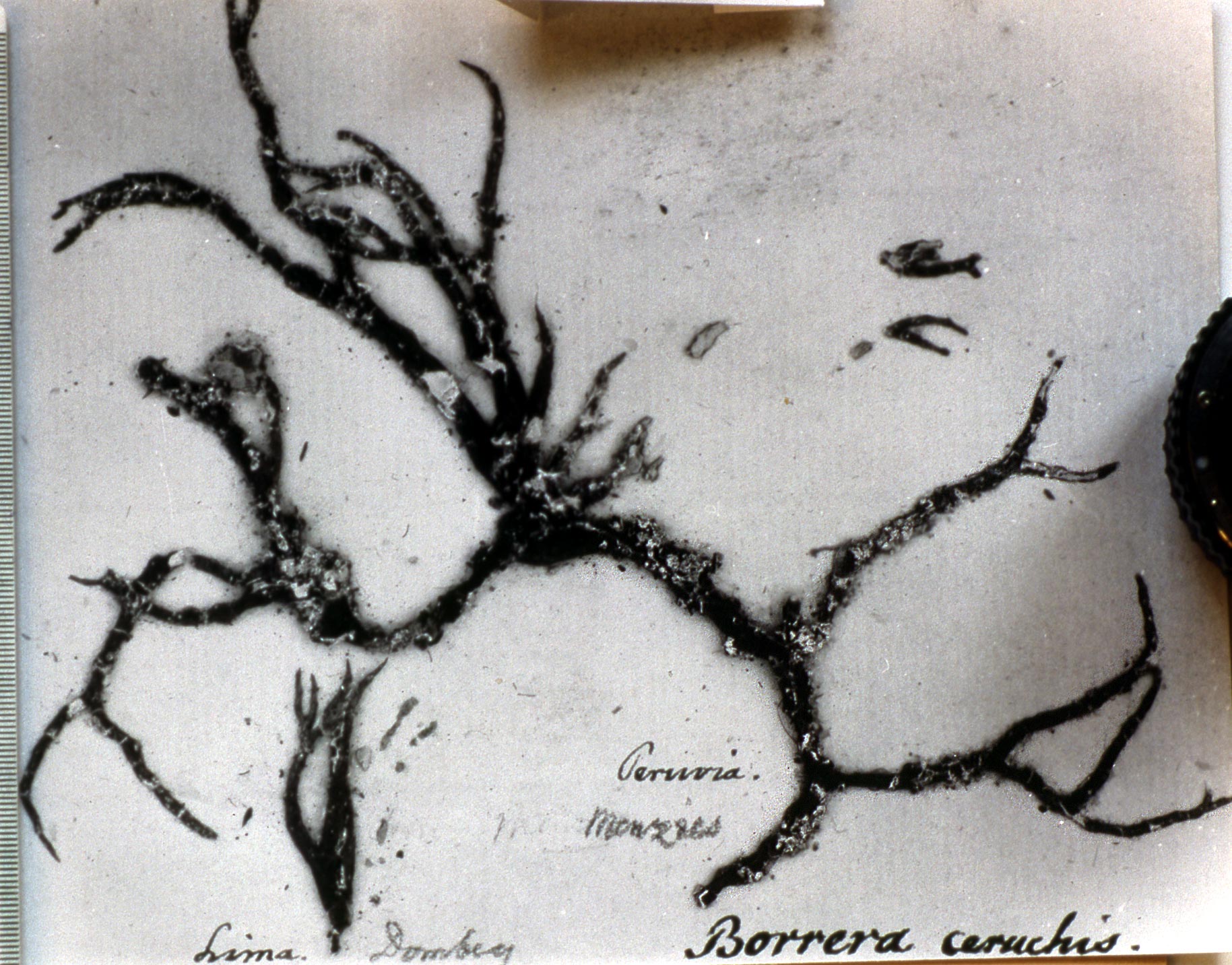
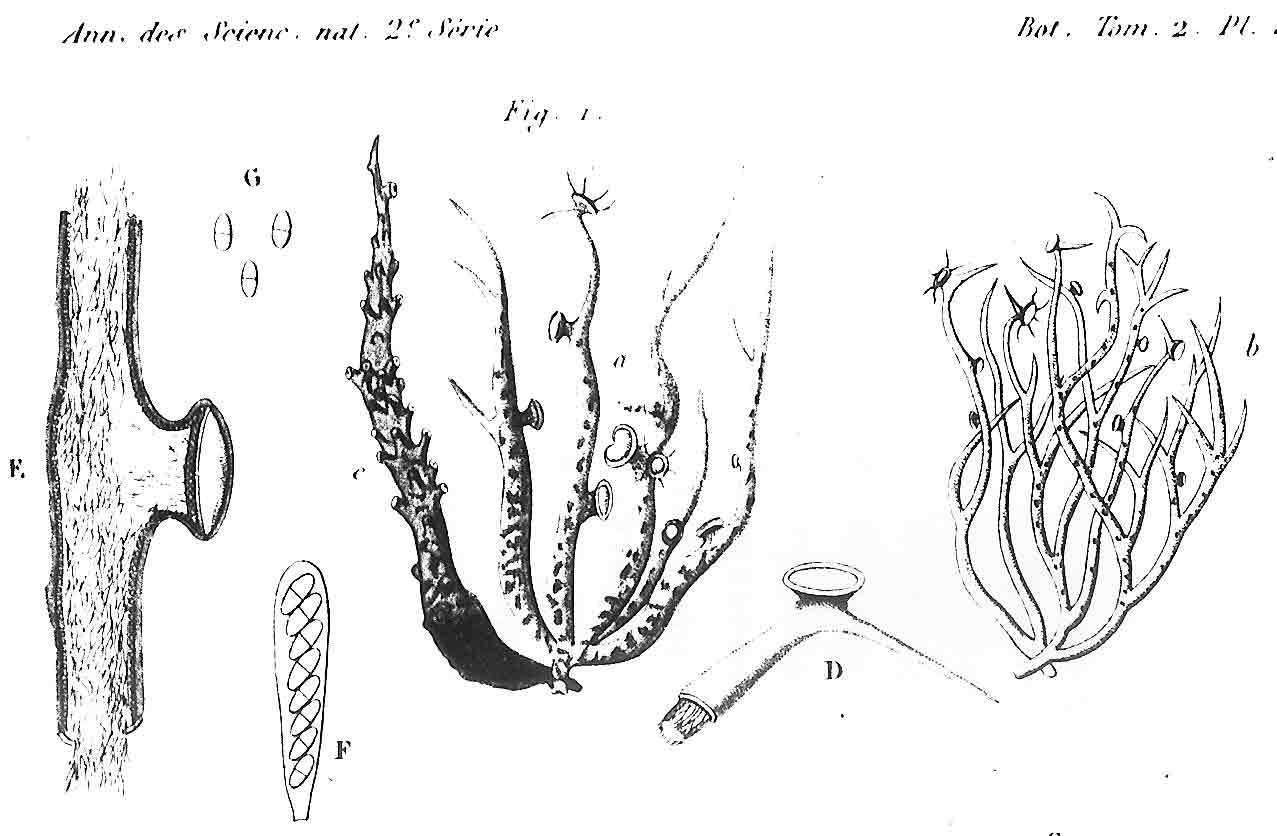
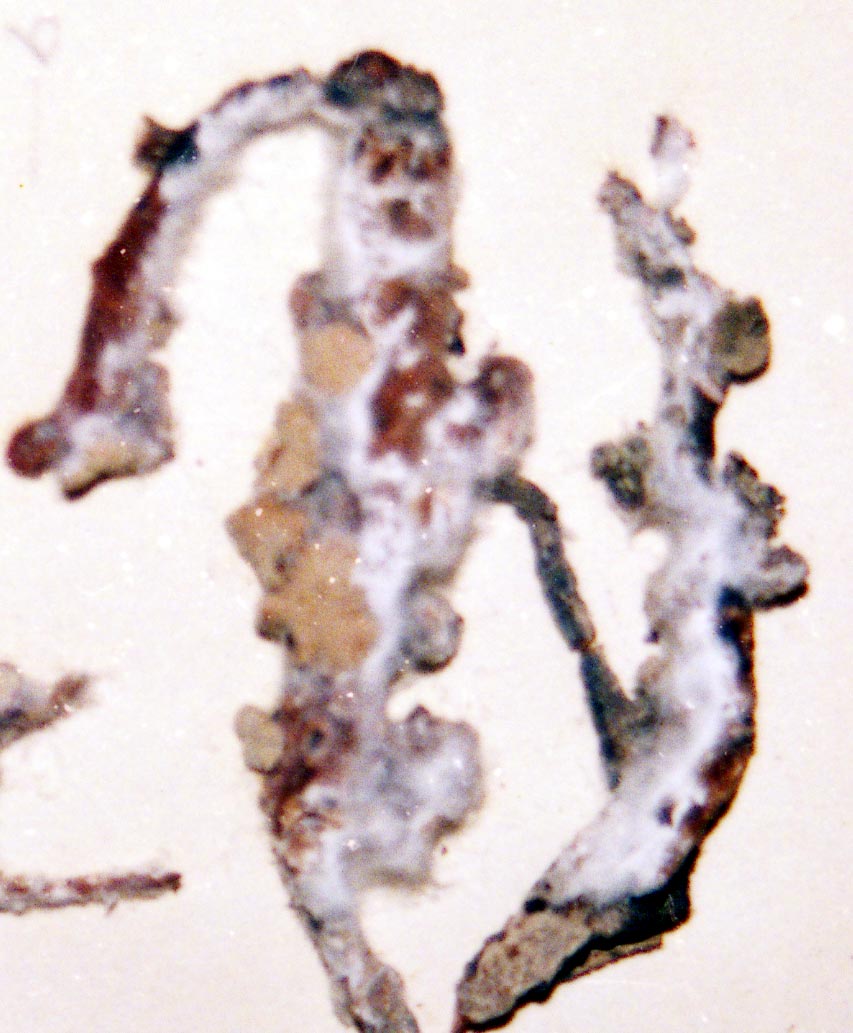
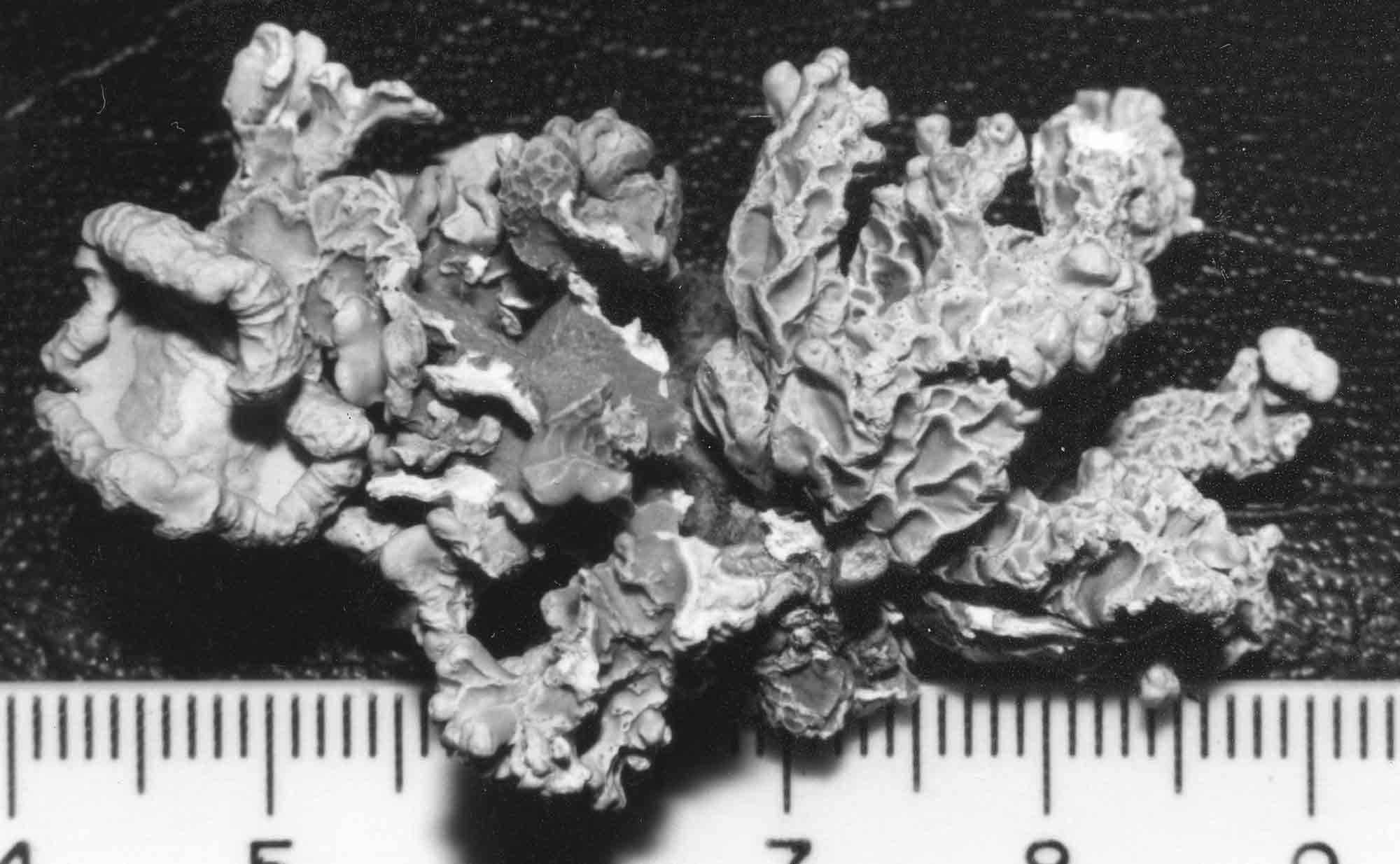
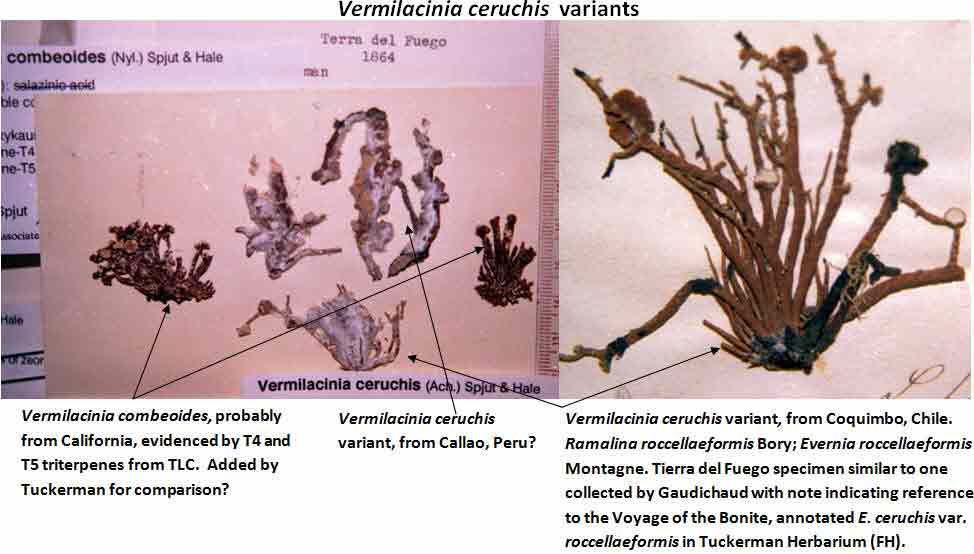
Several other variants of the Vermilacinia ceruchis complex in South America differ from their North American counterparts by lateral or subterminal apothecia, while also separated from the related South American species by the thallus branches united at base into a central attachment or holdfast, presumably as a result of growing on rocks. Subterminal apothecia are defined as those that develop ± solitarily on upper branches resulting in a change direction of growth from where they originate, or develop an appendage (spur) like branch (especially subgenus Cylindricaria). This in contrast to lateral multiple apothecia on a relatively straight branches as seen in the specimen from Callao. It is not known whether the Callao variant is extant, although it was reported by Gaudichaud to be abundant on rocks at Callao (Montagne 1834). The illustration immediately above is from Montagne (1834, as Usnea ceruchis). Except for the spine-like extension on apothecia, which it seems that Montagne may have added to make it appear more like Usnea, the center and left figures compare closely to the specimen above thought to be from Callao, Peru. The specimen on right that is more branched may have come from spines of cacti or branches of arborescent shrubs at Coquimbo, or possibly terricolous as it resembles V. tigrina from Chile, see image below from Follmann (1966). It is interesting to note that specimens in the Tuckerman Herbarium (Farlow Herbarium) shown above—mounted together on one sheet—correspond to Tuckerman's (1882) description and comments for Ramalina combeoides: “quite simple podetiiform thallus, and commonly terminal, now clustered apothecia, grows the the next species in California (Bolander [in reference to R. homalea, Nylander monograph]) but, though certainly marked, is inseparable from South American forms (Tierra del Fuego, Wilkes exp.).” Thus, Tuckerman (1882) regarded the North American V. combeoides to be the same as saxicolous variants of V. ceruchis, except for the terminal apothecia as he himself indicated. It should also be kept in mind that V. ceruchis and Niebla homalea were treated in separate genera by Acharius (1810), and that Montagne (1852) combined them into a single genus and species, only to be soon segregated (Trevisan 1861) and later classified into three groups (Bowler 1981, 'homalea', 'combeoides', and tigrina ['ceruchis' misapplied]), and two genera (Spjut 1995).
|
||
|
|
||
|
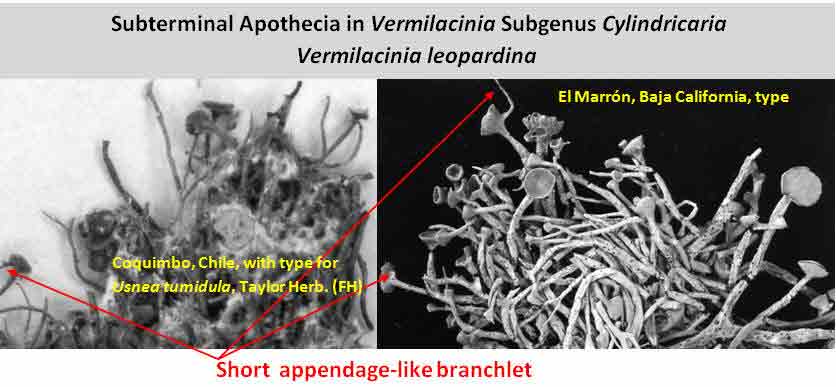
Subterminal
apothecia are seen more often in Subgenus Cylindricaria, usually
accompanied by a relatively short continuous branch that ascends sharply upwards
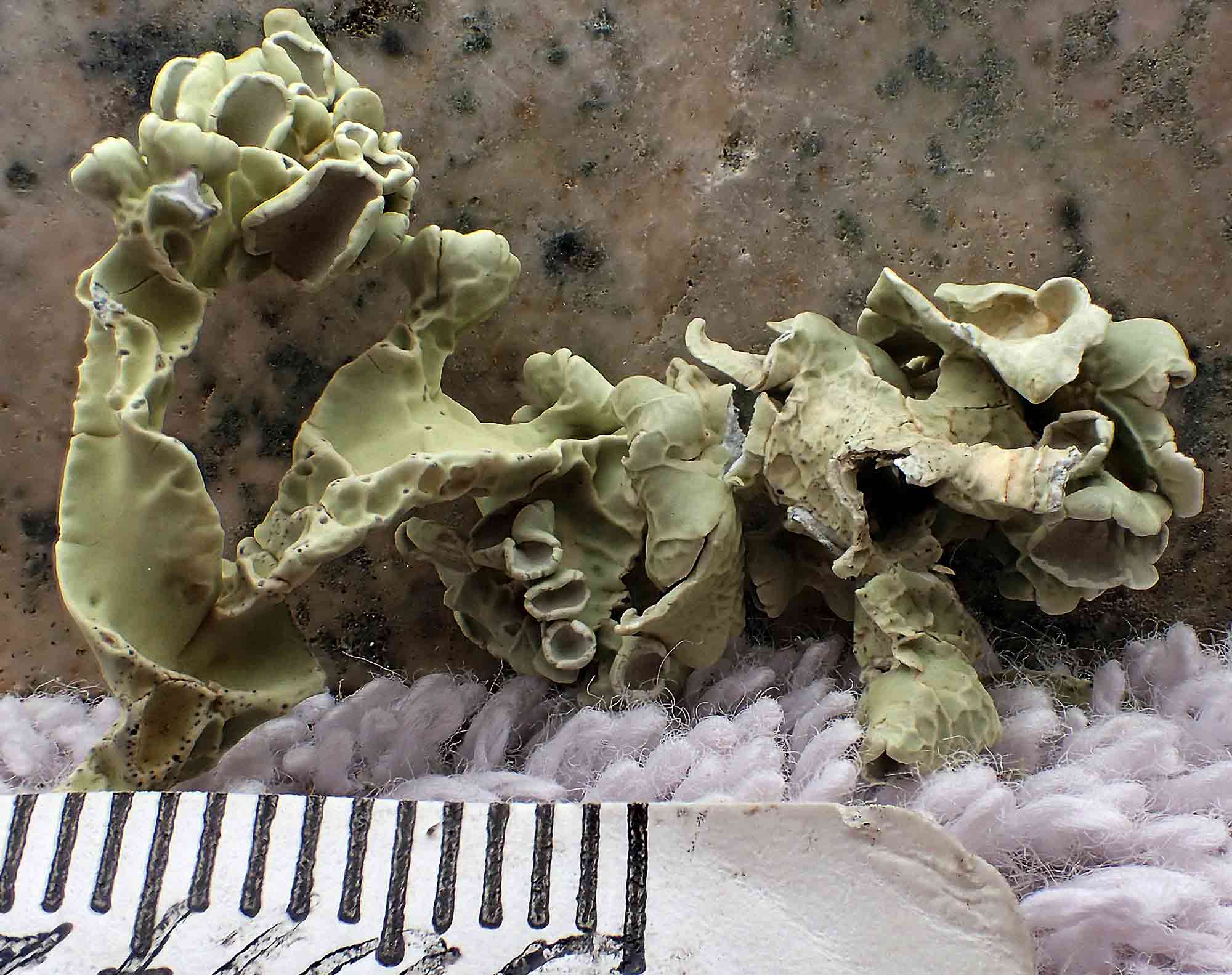
as shown above for V. leopardina.
This is in contrast to South American subgenus Vermilacinia
with
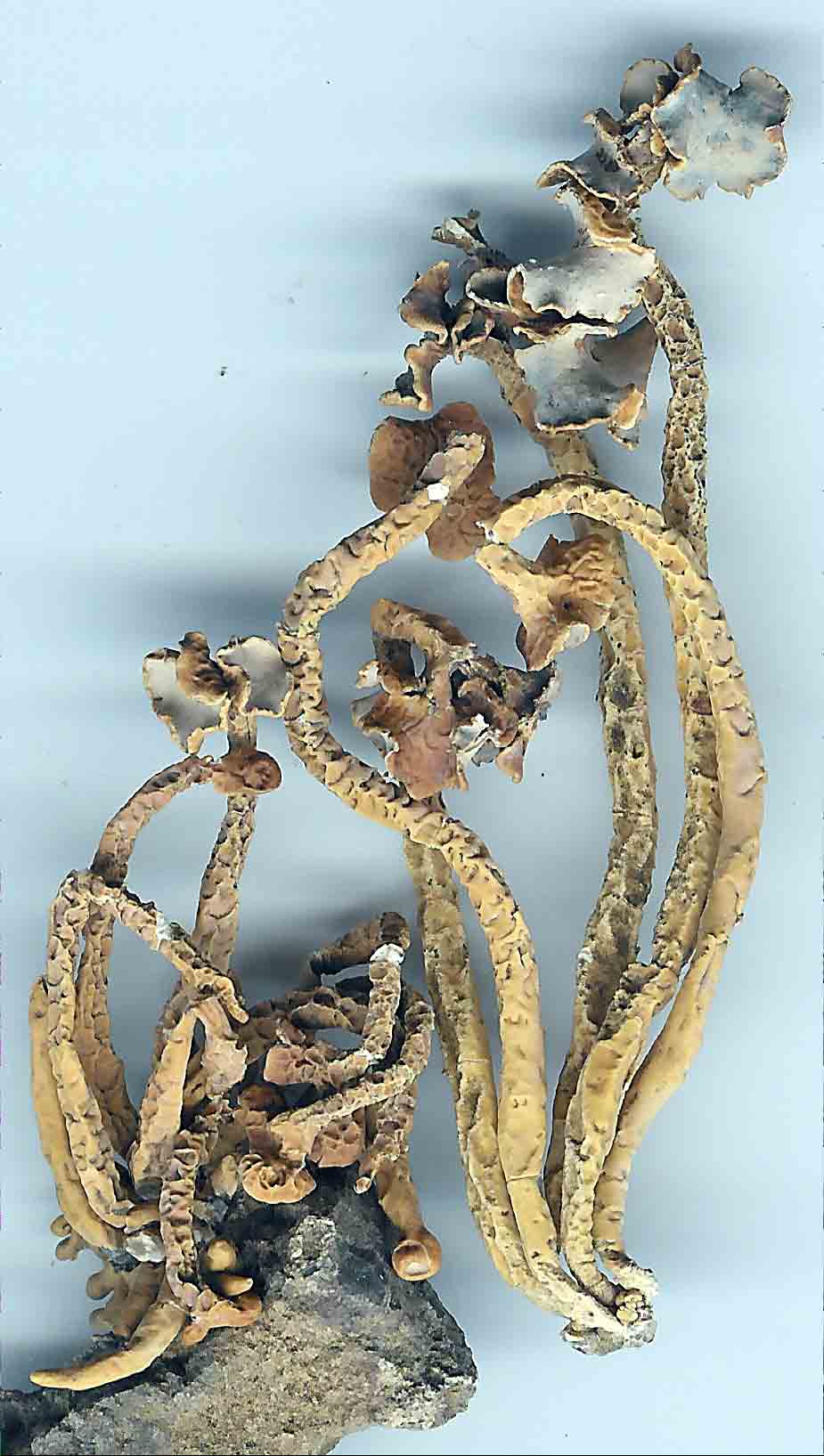
apothecial branches that bend abruptly (geniculate) as evident for the 'roccellaeformis
variant'.
The mildew-like appearance of the specimens is the result of efflorescence—a chemical change associated with the breakdown of the cortex from which the internal (medullary) hyphae erupt. The decorticating condition has been attributed to [-]-16 α-hydroxykaurane, a diterpene found in all species of subgenus Vermilacinia, most North American species of subgenus Cylindricaria, but not in Niebla (Spjut 1995, 1996). This diterpene is apparently rare in lichens, although known in mosses (Physcomitrium). The mold-like condition develops within a period of six months after a specimen has been collected; however, efflorescence can be prevented by storing specimens in a refrigerator below 40°F, although there may be a trade-off in that long term storage in a frost-free refrigerator causes the cortex to become brittle over time (gen. obs., ~9 yrs). Efflorescence is common in subgenus Vermilacinia and V. cephalota, V. cerebra, and V. tigrina in subgenus Cylindricaria, in which its occurrence seems closely associated with an unidentified triterpene (T3, Spjut 1996), and with bourgeanic acid, a distinct chemical class of aliphatic depsides (White & Johnson 1994). These compounds—and also the triterpene zeorin—occur in both subgenera but not in Niebla. Indeed, Niebla and Vermilacinia have little in common in their secondary metabolites except for occasional weak presence of accessory lichen depsidones in Vermilacinia. The distinction has been further sharpened by finding that species in the Old World 'Bourgeana clade'—also classified in Niebla (Rundel & Bowler 1978; Bowler & Marsh 2004)—are nested within the Old World Ramalina (Sérusiaux et al. 2010).
|
||
|
Summary: Key to South American species of Vermilacinia subgenus Vermilacinia
1. Basal branches solitary, or without a
central attachment point; apothecia lacking;
terricolous................................2
2. Thallus a
single cylindrical 'stem', 2–3 mm wide near base, sparingly branched;
Valparaiso and
3. Basal branches loosely united,
wide spreading, long attenuate; Coquimbo Chile,
extinct?.....................V. tumidula
4. Basal branches erect, inflated;
apothecia lateral; Callao Peru (see also V. vesiculosa, N
Am)..........Vermilacinia sp.
|
||
|
Subgenus Vermilacinia in North America
|
||
|
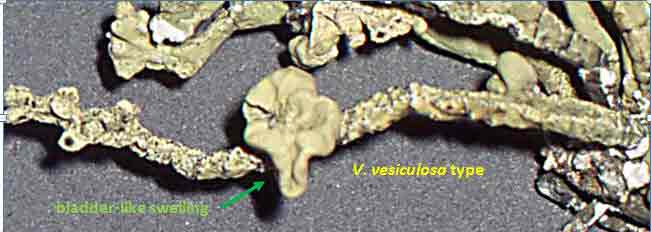
The North American species in the subgenus Vermilacinia are usually recognized by subterminal to terminal, often aggregate apothecia, in contrast to solitary to subterminal or lateral apothecia in the South American species. Included are species without apothecia found on rocks rather than on earth. One species, V. vesiculosa, is recognized for lateral large bladder-like (utricular) swellings and for the pycnidia that develop in tubercular lobes that form the thalline margin. Bladder-like or gall-like outgrowths ~ 1 mm—that are common on Vermilacinia and also Niebla—contain lichenicolous Heterobasidiomycetes Tremella nieblae (Diederich 2007); however, the development of lateral swellings in V. vesiculosa and the terminal swelling in the corticolous V. cerebra, appear related to an aborted development of apothecia. The utricular swellings in V. vesiculosa are exceptionally large, ~10 mm diam. Four species—Vermilacinia johncassadyi, V. ligulata, V. reptilioderma, and V. rosei—are recognized by triterpenes in RF Classes 1–2—than what characterizes most species in Subgenus Vermilacinia. The species occur in southern Baja California, Isla Cedros, and on the Vizcaíno Peninsula and nearby Isla San Roque. In TLC, these terpenes appear quite strong, and seem to replace terpenes in higher RF classes, but not zeorin and [-]-16 α-hydroxykaurane. Vermilacinia johncassadyi is morphologically similar to V. laevigata. Vermilacinia ligulata may be compared to V. polymorpha in the Channel Islands in having relatively short basal branches; the Baja California species is ± digitately divided into twisted lobes, whereas the California species has more cylindrical dichotomously divided branches. Vermilacinia reptilioderma, a relatively rare species, is closely related to V. paleoderma, a common species along the Pacific Coast south of Punta Baja. They are sympatric on the Vizcaíno Peninsula and on Isla Cedros and can only be distinguished with confidence by their lichen substances. Similarly, V. rosei strongly resembles V. varicosa, in which they are sympatric on Isla San Roque, and V. rosei has also been collected on Isla Cedros and recently on the Vizcaíno Peninsula. Except for V. ligulata, they may be regarded as sibling species. The following key defines the species in the North American Vermilacinia subgenus Vermilacinia. |
||
 |
||
|
Key to
Species of Subgenus Vermilacinia: North America
1 Cortical surface
with regular protruding ridges, creases, or crater-like depressions…........................................… 2
2 Thallus cushion-like, of short branches
tightly compacted, 1–2 (-4) cm high, rarely unbranched for more than
3(2) Thallus of mostly
undivided tubular branches
with terminal or subterminal apothecia, the branches arising from
3 Basal branches simple to shortly bifurcate near
apex, or densely branched below apex, often matted
4 (3) Thallus lacking terminal acicular branchlets, or abruptly with
narrow acicular isidia; basal branches
sparingly
5(3) Basal branches closely compacted, without isidia or soredia, mostly simple
and stubby, or shortly divided
7(2) Branches with bladder-like swellings; pycnidia elevated on pustular
lobes; rare, between
8(7) Primary branches 5–10˟ longer than wide, twisted, lobulate; triterpenes present in RF classes 1–2; Isla
9(8).
Branches flexuous, 0.5–1.5 mm wide; cortex on upper branches thinner,
eroded and pitted;
10(9).
Apothecia aggregate on expanded marginal and terminal lobes constricted below;
endemic to
11 Triterpenes present in RF Classes 1–2;
Vizcaíno Peninsula and Isla
Cedros (V. lacunosa if
12(1) Branches distinctly blade-like, two-edged.........................................................................................................
13
13(12) Triterpenes present in RF
classes G: 1–2; rare, Isla Cedros, southern half of
14(13) Branches uniformly blackened around base to slightly above,
usually less than 4 cm long,
1–4 mm
14(13) Branches
irregularly blackened near base and above, usually >3.5 cm long and 4–7
mm
wide;
15(13)
Branches uniformly cylindrical, irregularly blackened above base, often with
black transverse bands or cracks........
16
16(15)
Apothecia mostly terminal; Californian coastal chaparral region, San Francisco to San Quintín,
17(15)
Branches dilated, flattened and/or digitately divided;
Baja California.................................................................... 18
18(17) Triterpenes present in RF Classes 1–2; Isla San Roque
and Isla Cedros...................……..……... Vermilacinia rosei
19(17)
Branches inflated, ± round in x-section; Isla Guadalupe, Channel Islands, peninsular
BCN from
|
||
|
Additional images obtained from review of specimens collected for molecular
|
||