|
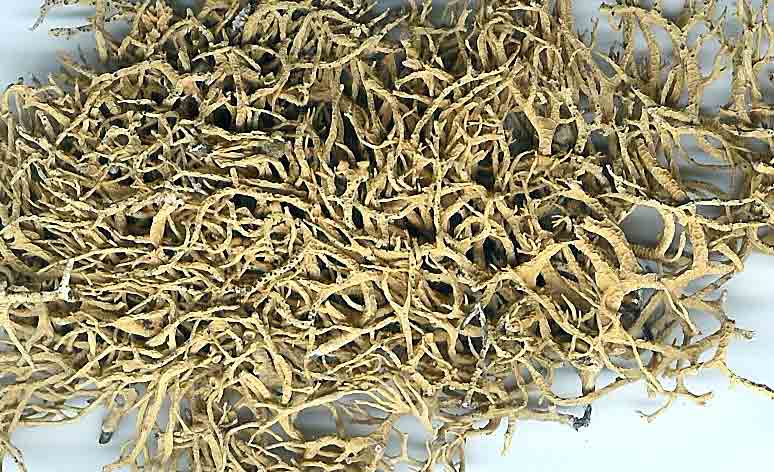
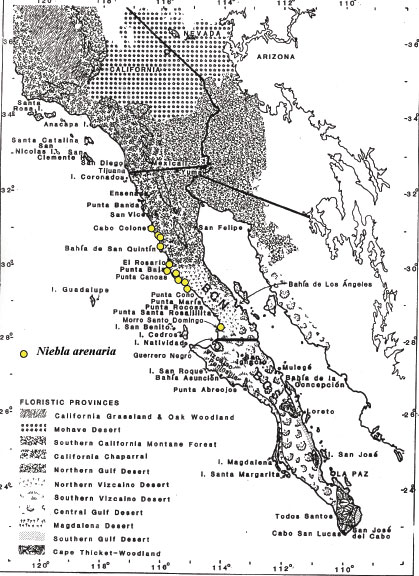
Niebla arenaria is a fruticose lichen endemic to Baja California,
ranging from its southernmost occurrences near Morro Santo Domingo north to near Colonet
(Spjut 1996). It often occurs loosely attached to soil on barren sandy beaches along the Pacific Coast
extending slightly inland
on alkaline clay or gravel among low scrub vegetation, often with
Frankenia palmeri and
Atriplex julacea, or with
Ambrosia
spp.,
Euphorbia misera, and various cacti.
Niebla arenaria
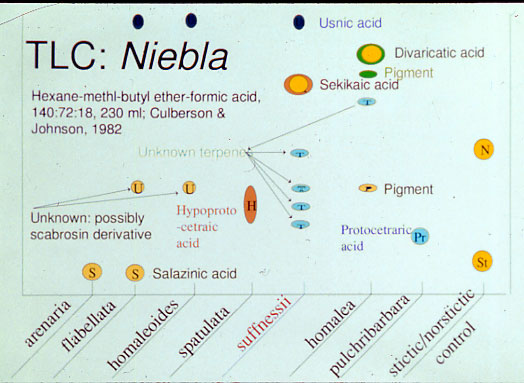
is distinguished by its chemistry of salazinic acid, which may be accompanied by
consalazinic acid, or trace of protocetraric acid, and by its morphology of a hemispherical thallus without a
holdfast, intricately divided into
numerous narrow ribbon-like branches, appearing antler-like and shortly
bifurcate near apex. Black pycnidia
are usually prominent at the tips of shortly bifurcate branchlets. Thalli
collected near Punta Baja have longer terminal branches abruptly bifurcated near
apex.
Niebla arenaria is often gregarious, presumably from reproducing
asexually by fragmentation of terminal bifurcate branchlets. It can form a single dominant Niebla
community on sandy substrates as observed along the northern shore Bahía Falsa,
the type locality, in May 1986. Thousands if not millions of thalli are recalled
to have covered the shoreline, the cover measured by Rundel et al. (1972) at
70%. However no thalli were seen at type locality in January 2016, although some
were encountered ~ 500 m south on less disturbed beaches.
Niebla arenaria can also be found in mixed Niebla terricolous
communities with
N. effusa,
N. josecuervoi, and
N.
juncosa on mesas southwest of El Rosario to Punta Baja.
It appears most common in the chaparral-desert transitional region.
Other species with terminal
antler-like branchlets are
Niebla limicola
and
Niebla brachyura.
Niebla limicola differs by having flattened and expanded branch parts
from which the short bifurcate branchlets develop. It is most common south of
Morro Santo Domingo.
Niebla brachyura differs by having hyprotocetraric
acid instead of salazinic acid; it is
relatively infrequent, found mainly in the southern region of the Northern Vizcaíno
Desert, and on Isla Cedros.
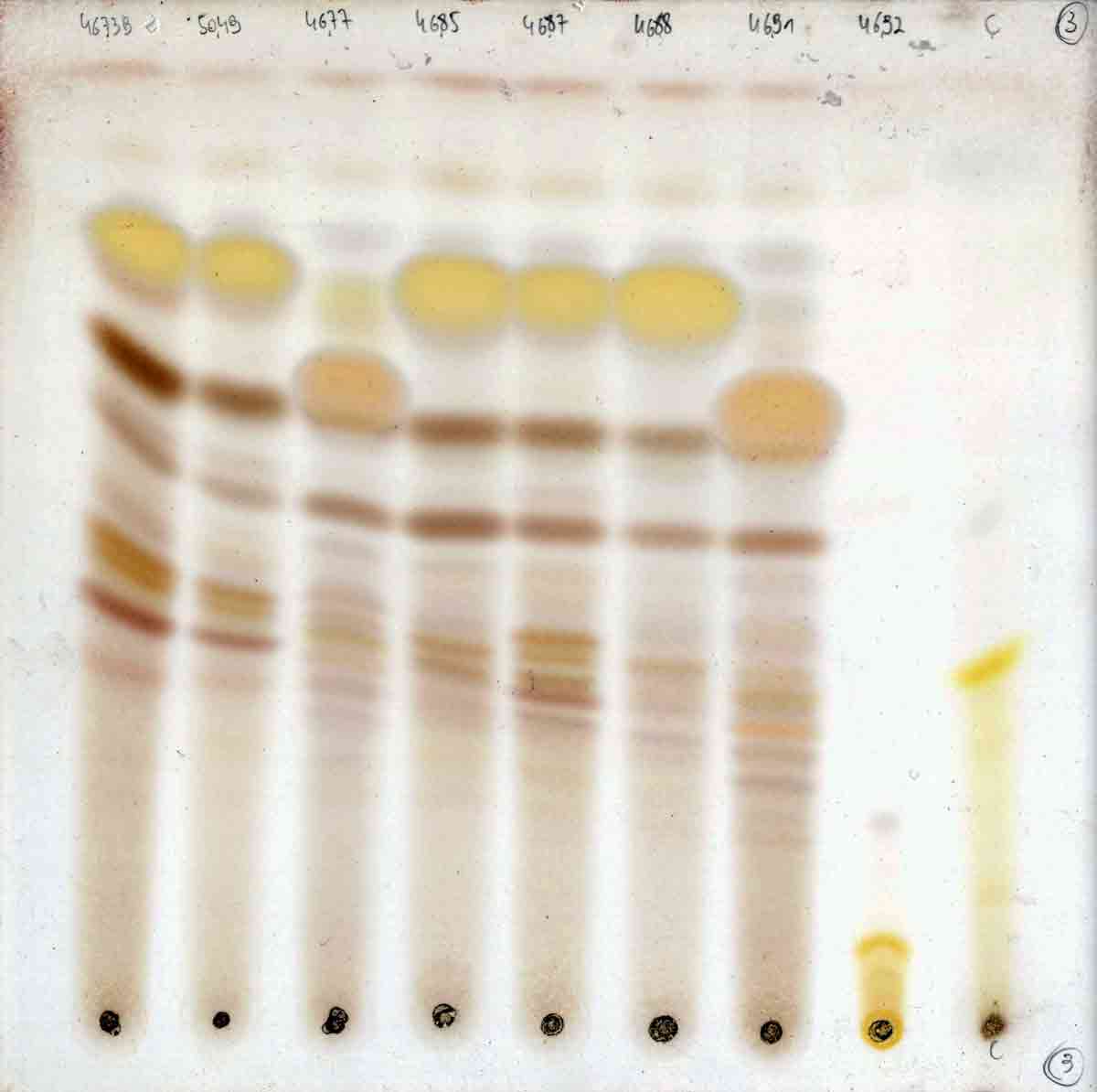
The geographical
distribution of Niebla arenaria shown on the map of Baja California is
based on more than 100
specimens analyzed by thin-layer chromatography (TLC). The type collection (Spjut,
Cassady & McCloud 9327, holotype at US) from Bahía Falsa near San Quintín)
included perhaps another 100 thalli that were extracted and tested for
anticancer activity. In addition to isotypes distributed to institutions
in Baja California (BCMEX) and California (LA), other specimens were
submitted to the lichen exchange in Arizona (ASU).
Although Niebla arenaria can be recognized as described by Spjut (1996),
its phylogeny is not resolved (Spjut et al. 2020) along with most species that
lack triterpenes that are segregated by their depsidones (absent
in N. homaleoides),
and by their by branch morphology (flat vs. tubular-prismatic, irregularly
widened vs. sublinear) and their terminal branching (antler-like or terminally
dilated and fringed). Two accessions in Spjut et al. (2020, Fig. 7) are
shown in different clades; both Stacey and BPP analyses suggest they represent
two species, one collected approximately 500 m from type locality in the
Chaparral-Desert Transition near San Quintín, the other from the southernmost
region of the Northern Vizcaíno Desert, the type locality for N. marinii.
Additionally, the type for N. arenaria was collected in the vicinity of the type locality
for N. pulchribarbara, a sandy bay shore along Bahía Falsa. The latter
distinguished by having protocetraric acid (Rundel et al. 1972; Spjut 1996), and
by the lack of short bifurcate branching near
apex. The type for another related species, N. josecuervoi, which also
has salazinic acid, was collected along the same peninsula on a
ridge of volcanic rocks. Its branches are tubular-prismatic appearing
mostly linear throughout, while often bearing short comb-like branchlets along
primary branches; the comb-like branches often much longer than the terminal
bifurcations near apex, in contrast to just the relatively short bifurcate
branchlets in N. arenaria and N. limicola.
Additional References: See
Niebla.
| 


















{kind=link}
{kind=link}
{kind=link}
{kind=link}