Niebla brachyura is
a fruticose lichen that has been recognized as endemic to the Northern Vizcaíno Desert of Baja California
from near El Rosario south to Morro Santo Domingo (Spjut 1996). It is distinguished
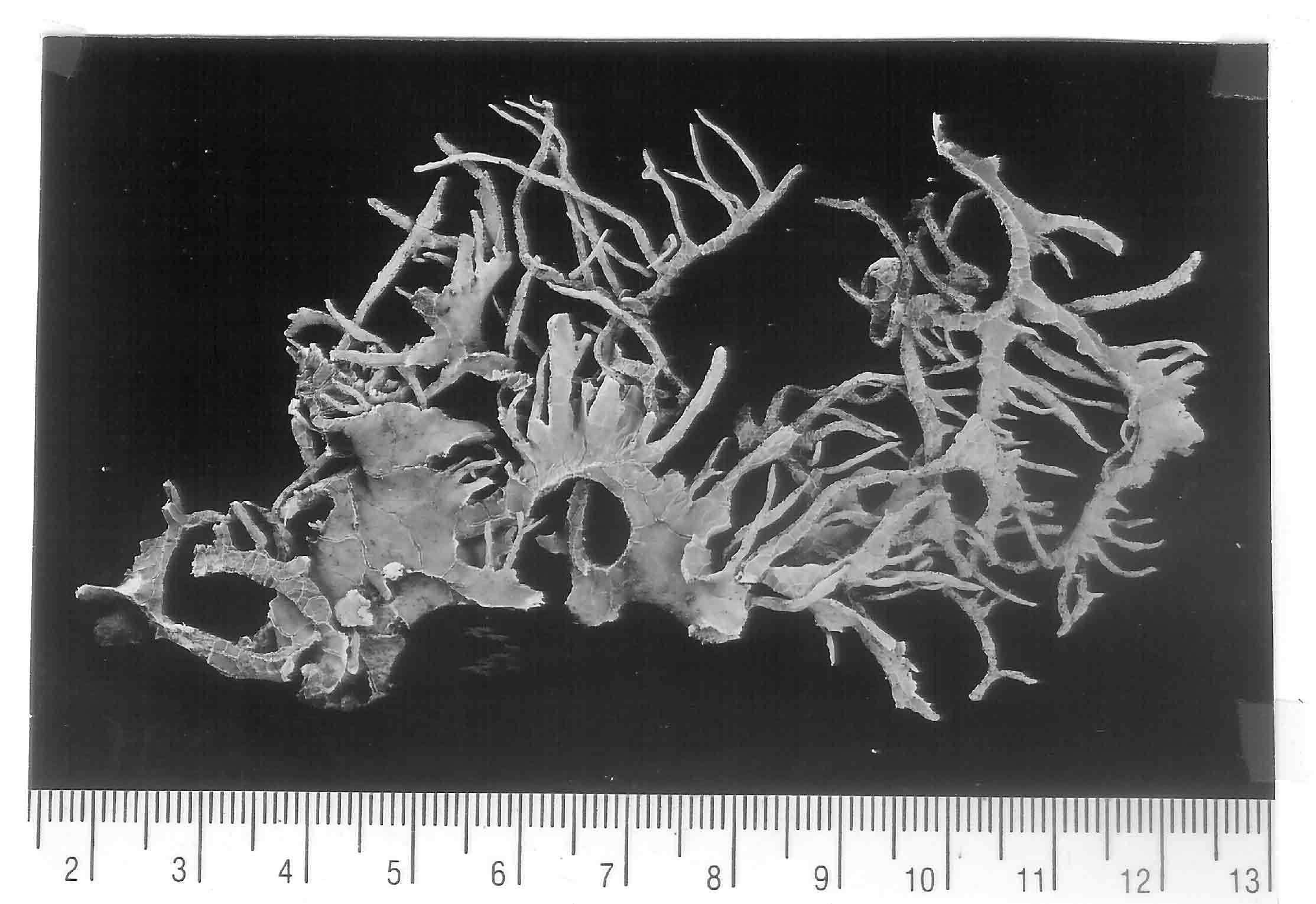
by its hemispherical thallus intricately divided into narrow to irregularly
widened branches that are often shortly bifurcate near apex, and by its
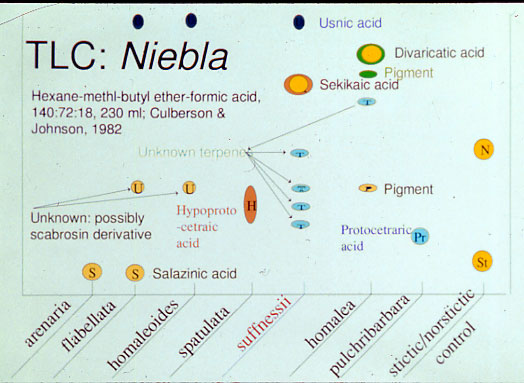
lichen substance of hypoprotocetraric acid. Niebla brachyura is one of two species
in the genus that was recognized by hypoprotocetraric acid (Spjut 1996);
however, specimens collected in 2016 for Spjut et al. (2020) discovered
hypoprotocetraric acid in combination with sekikaic acid or protocetraric acid. The
other, the N. spatulata complex, occurs in the
Southern Vizcaíno desert, from near El Tomatal,
west across the Vizcaíno
Peninsula, to southern Isla Cedros; it often includes demethylnotatic acid and
conhypoprotocetraric acid, while closely associated with thalli containing
salazinic acid.
DNA phylogeny
for references cited above
indicate that
hypoprotocetraric acid does not alone distinguish the species in the Northern
Vizcaíno Desert; instead two or three species of Niebla, while one
specimen identified N. brachyura also has salazinic acid in
addition to hyproprotcetraric acid (Spjut et
al. 2020, Supp. file S5 ).
The type collection is a single specimen (Spjut & Marin 9073K,
holotype) taken from a ~ 200 g sample of mixed species collected in May 1985 for
the National Cancer Institute anticancer/antiHIV screening. Specimens in
the sample analyzed by TLC were found to have either
divaricatic acid, identified
N. caespitosa, N. eburnea and
N. flagelliforma or
salazinic acid,
N. flabellata. Consequently, the sample
was retained entirely for taxonomic study. The type for N. flabellata was also selected from this sample
(Spjut & Marin 9073k). It was not until five years later that Niebla
brachyura was found at other sites, one near Punta Canoas where six
specimens were collected in association with N. arenaria,
then
further south at
Punta Cono with other species of Niebla and also Vermilacinia, and
recently, in Feb 2016, at Morro Santo Domingo.
Niebla brachyura
is phenotypically similar to salazinic-acid species, N. arenaria, N. limicola and
N. effusa Although N. brachyura
is easily
distinguished by its chemistry of hypoprotocetraric acid, this distinction is
not entirely supported. Four
specimens cited in Spjut et al. (2020) did not collectively occur in a distinct clade;
rather, they were distributed in four different microclades, three were collected
at Morro Santo Domingo, a fourth along road from El Rosario to Punta Baja (Spjut et al. 2020).
Species delimitation by BBP recognized three species, Stacey 2 species, where
Spjut (1996) recognized only one. Thus, two of the DNA species would
have to be from the same locality, Morro Santo Domingo, while the third could
have come from there, or from near El Rosario. What distinguishes the
species other than DNA is cryptic. One of the Morro Santo Domingo specimens,
17244, has salazinic acid in addition to hypoprotocetraric acid. 17262
might be distinguished by pycnidia immersed within the cortex but again their differences are not
evident in the phylogeny (Spjut et al. 2020, Fig. 7). Only one specimen
was included in Jorna et al. (2021, Fig. 5), 16-937, in a clade with N. marinii
and N. cf. limicola.
For more discussion
and reference materials see Introduction to Niebla